
| 
|
|
|
|
последующие испытания, опять-таки не зная о сделанном двумя
другими.
Все это тонкая работа, но при необходимых навыках за рабочий день можно прооперировать до дюжины цыплят. Они хорошо переносили операцию и наркоз: придя в себя, цыплята с поврежденным IMHV или LPO выглядели совершенно нормальными и не отличались ни от ложно оперированных, ни от интактных контрольных особей. После оценки поведения цыплят забивали, а поврежденные участки мозга исследовали под микроскопом. Как и в других случаях, для получения статистически достоверных результатов эксперимент многократно повторяли, чтобы в каждой опытной (не контрольной) группе насчитывалось двенадцать или более цыплят.
При четырех рабочих днях в неделю (это дни, когда вылупляются цыплята) с учетом времени, затрачиваемого на ложные операции и на проверку локализации повреждений, для получения одной серии результатов требуются примерно три недели; практически же на это уходит около месяца, поскольку что-то всегда идет не так: то цыплята не вылупляются, то нужно идти на какое-то заседание и т. п.1.
Потратив нужное число месяцев и сделав достаточно ошибок, чтобы освоить методику повреждения мозга, мы смогли приступить к первому этапу к проверке результатов Сэри. В начале 1989 года мы подтвердили их. Сомнений быть не могло:
1 Я давно уже на собственном печальном опыте убедился, что при планировании эксперимента нужно взять максимальное потребное время, удвоить его и добавить еще немного тогда ваши расчеты окажутся более или менее верными. Следует учитывать еще один важнейший момент: если вы налаживаете метод, которым раньше не пользовались, а позаимствовали из чьей-то статьи, он обычно начинает работать не раньше чем с третьей попытки; в двух первых вы каким-то непонятным образом обретаете в кончиках пальцев ту магическую, не поддающуюся описанию силу ("безмолвное знание", как назвал ее Майкл Полани, некогда химик, а потом философ), которая позволяет освоить метод не только в теории, но и на практике.
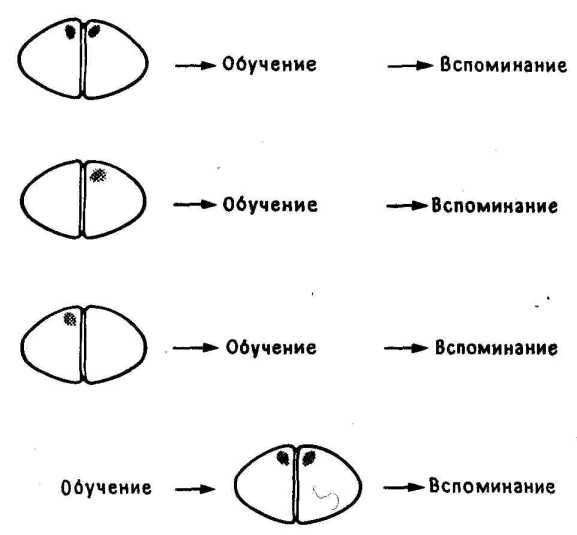
11.1. Повреждение IMNV. На этом и следующих рисунках затемненные участки места повреждения.
цыплята с поврежденным IMHV явно обучались пассивному избеганию, т. е. клевали горькую хромированную бусину, так же, как их ложно оперированные собратья, трясли головами, ощутив ее вкус, и отворачивались при повторном предъявлении. Но спустя несколько часов при вторичном испытании они полностью забывали приобретенный опыт и клевали сухую бусину столь же энергично, как и контрольные цыплята, которым раньше давали бусину, смоченную водой. Таким образом, повреждение мозга не влияло на поведение птенцов (клевание бусины), на чувство вкуса или общую подвижность они лишь не помнили, что следует избегать бусин определенного вида.
Теперь нужно было пойти дальше Сэри и производить одностороннее повреждение. И снова мы получили ожидавшийся результат. У цыплят с поврежденным левым IMHV реакция избегания отсутствовала, а при правостороннем повреждении полностью сохранялась.
Все было нормально в полном соответствии с прогнозом, основанным на ранее полученных данных, и с пятым критерием, для запоминания необходим был левый, но не правый IMHV ( 11.1).
По логике нашего подхода следовало теперь проверить, что
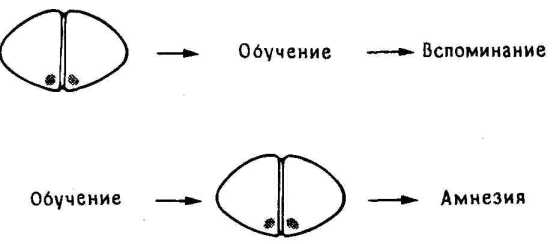
11.2. Повреждение LPO.
произойдет, если повредить мозг после обучения. Это можно было сделать не раньше чем через час, так как иначе на результатах испытаний могло бы сказаться действие наркоза. К нашему удивлению, даже двустороннее повреждение мозга спустя час после обучения не приводило к амнезии.
Итак, первый парадокс: для запоминания необходим интактный левый IMHV, но коль скоро цыплята уже усвоили задачу (по крайней мере в первый час после обучения), IMHV оказывался ненужным [2].
Куда же переместилась память? Учитывая все полученные ранее биохимические и морфологические данные, следовало искать ее следы в LPO. Эксперимент подтвердил это. Двустороннее повреждение LPO через час после обучения действительно вызывало амнезию, которой не было при одностороннем правом или левом повреждении. По-видимому, в норме след памяти после обучения каким-то путем мигрирует из IMHV в LPO, чем и объясняется отсутствие амнестического эффекта повреждения после тренировки.
Это был весьма интересный результат, так как он позволял понять, почему мы находим биохимические и морфологические изменения и в IMHV, и в LPO. Тот факт, что для сохранения памяти достаточно одного LPO, правого или левого (одностороннее повреждение не вызывало амнезии!), согласовался с тем, что многие из выявленных нами изменений имели место как в правом, так и в левом LPO, т. е. след памяти о бусине и реакции избегания сохранялся в обоих полушариях ( 11.2).
Не довольствуясь столь простым результатом, мы пошли дальше. Что будет, если повредить LPO до обучения? К нашему удивлению, такая операция никак не влияла на память [3] ( 11.2).
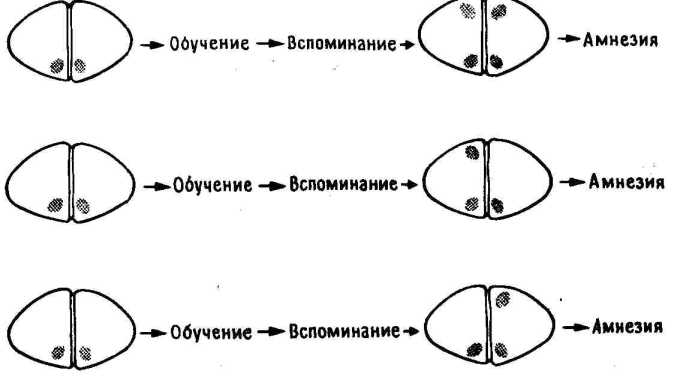
11.3. Повреждение LPO до обучения и LMHV после него.
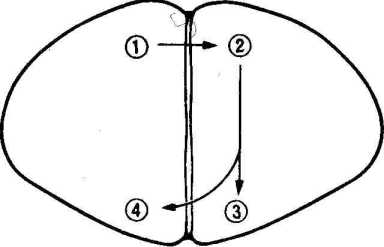
11.4. Объяснение результатов: возможные перемещения следов памяти.
Если пассивное избегание вырабатывается и при отсутствии LPO, то не происходит ли реорганизации следа памяти каким-то иным образом? Может быть, в этом случае след просто закрепляется в IMHV? Если это так, то повреждение LPO до тренировки и повреждение IMHV после тренировки две операции, по отдельности не приводящие к амнезии, при их сочетании у одного и того же цыпленка должны будут вызвать амнезию. Так оно и оказалось в действительности ( 11.3). Получив такие результаты, я предположил, что решающая роль принадлежит IMHV.
Но Дейв думал иначе. Тогда мы произвели односторонние повреждения, и выяснилось, что он прав. У цыплят, не имевших LPO во время обучения, последующее; повреждение левого IMHV не влияло на запоминание, а повреждение правого IMHV вызывало амнезию. .,Все это схематически показано на 11.3.
Результаты были интригующими. Размышляя над ними, я решил, что (все можно объединить, исходя из несколько механистической модели, в которой первичный след памяти о бусине и реакции избегания образуется в левом IMHV, а затем, спустя несколько часов после обучения, "перекочевывает" сначала в правый IMHV, а потом в правый и левый LPO ( 11.4).
Я пришел к этой схеме исключительно на основании логики экспериментов с повреждением мозга. Но подобная схема шокирует нейроанатомов, у которых тут же возникает очевидный вопрос: разве есть какие-то прямые нервные пути между IMHV и LPO? Согласно простой концепции коннекционизма, такие пути действительно необходимы. В то время, когда мы приступали к исследованию, мы не знали, существуют ли они, но это казалось маловероятным. Нам было известно, что все нервные пути от органов чувств, таких как глаза и вкусовые рецепторы, сходятся к IMHV.
Мы знали даже о непрямых связях между IMHV и LPO. Именно поэтому IMHV рассматривались как некие "ассоциативные зоны", где интегрируется разнообразная входная информация например, сопоставляется зрительное восприятие бусины с ощущением ее вкуса. Напротив, LPO находятся на выходной стороне мозга, в области, ответственной за двигательные функции (как, например, клевание) и, возможно, также за реакции "эмоционального" типа. После начала наших опытов анатомия мозга цыплят стала несколько яснее.
Между IMHV и LPO действительно нет простых связей, хотя, по-видимому, существует непрямой путь через третий отдел мозга так называемый архистриатум.
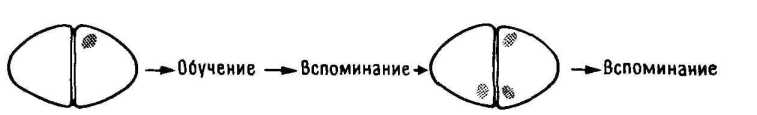
11.5. Блокирование перемещений.
Но и без учета анатомических данных приведенная гипотетическая схема ( 11.4) перемещения следов памяти позволяет сделать ряд предсказаний. В частности, из нее следует, что повреждение правого IMHV до начала обучения (само по себе не вызывающее амнезии) должно прерывать последовательность событий, и тогда след памяти "застрянет" в левом