
| 
|
|
|
|
IMHV, не имея возможности переместиться в LPO. Повреждение после обучения, обычно приводящее к амнезии, теперь уже не вызовет ее. Высказав это предположение, мы тут же (во всяком случае не позднее чем через месяц) проверили его ( 11.5).
Оно подтвердилось. Тогда Дейв, Терри и я решили опубликовать обобщающую статью с описанием всех новых экспериментов. Мы быстро подготовили черновой вариант, но, когда уселись, чтобы вместе доработать его, кто-то (сейчас уже не помню кто) неожиданно сказал: "А что, если до обучения повредить правый IMHV?".
Мы знали, что само по себе такое повреждение не дает амнестического эффекта, а наша модель предсказывает, что след памяти в этом случае должен оставаться в левом IMHV просто из-за отсутствия выхода. Поэтому у цыплят с поврежденным до обучения правым IMHV повреждение левого IMHV после обучения приведет к амнезии. Такой вариант опыта мог действительно стать окончательной проверкой.
Мы отложили статью в сторону и потратили еще месяц на новый эксперимент. Вот что мы получили ( 11.6):
11.6. Но след находит другой путь...
До этого я был готов, как некий научный Пуаро, собрать всех подозреваемых в гостиной и указать виновного, как было совершено преступление, то бишь как работает память. Теперь это было исключено. Эксперимент дал однозначный результат целиком отрицательный. Ничего, пустота, нуль, никакой амнезии. След памяти не мог перейти в правый IMHV, но он определенно не остался и в левом.
Куда же он подевался?
Почти полвека назад психолог Карл Лэшли написал классическую статью, обобщив результаты десятилетних экспериментов с обучением крыс. Он обучал их ориентироваться в сложных лабиринтах, а потом удалял отдельные участки мозговой коры, чтобы выяснить, где сохраняются следы памяти. К своему удивлению, он не обнаружил специфических участков, удаление которых приводило бы к полной утрате усвоенного навыка; вместо этого навык постепенно ухудшался по мере увеличения размеров удаленной области коры.
Создавалось впечатление, что память зависит просто от количества мозгового вещества. На основе этих наблюдений Лэшли построил кон-
цепцию "эквипотенциальное™" коры. Свою статью он озаглавил "В поисках энграммы" и закончил ее выводом, что память одновременно присутствует в мозгу везде и нигде [4].
Опыты Лэшли и его неутешительные выводы утратили актуальность в результате последующих экспериментов, однако парадокс локализации памяти остался. Это напоминает мне, во-первых, о том, что при повреждении IMHV и LPO мы изучаем не функции этих областей, а функции остального мозга, которые быстро перестраиваются в отсутствие двух первых; и, во-вторых, о том, что память нельзя понимать как нечто статичное, "находящееся" строго в одном месте или в небольшой группе клеток; она существует в более динамичной и рассеянной форме. Кроме того, мозг это тонко и сложно организованная структура с многообразными средствами функционального обеспечения. Вы мешаете ему осуществлять какую-то деятельность, а он находит другие пути к той же цели. Блокируете оптимальные связи между IMHV и LPO, a цыпленок находит им замену.
Мозг действует не как набор стандартно соединенных мелких ячеек, а как функциональная система, насыщенная разнообразными связями. Конечно, всегда следует помнить об анатомии мозга, но нельзя замыкаться в ней, поскольку ясно, что и сам мозг не сводится к анатомически выявляемым структурам. Настаивать на его механистическом объяснении, игнорируя мои предостережения о неразумности редукционизма, это значит обрекать эксперимент на погружение в трясину парадоксальных результатов.
Двойные волны
Мысль о том, что у нормальных цыплят при обучении возникает своего рода поток запоминаемой информации из левого IMHV в LPO, приводит к еще одному парадоксу и подсказывает новые эксперименты. Вся совокупность накопленных за десятилетие биохимических и морфологических сведений, рассмотренных в предыдущей главе, сводится к тому, что у цыплят, обучавшихся избегать горькую бусину, происходят долговременные изменения в химии и структуре клеток LPO и левого IMHV. Однако опыты с повреждением мозга, по-видимому, показали, что обученному цыпленку IMHV вовсе не нужен.
Почему же тогда сохраняются изменения в этой области?
Я размышлял об этом по вечерам, а днем пытался заполнить
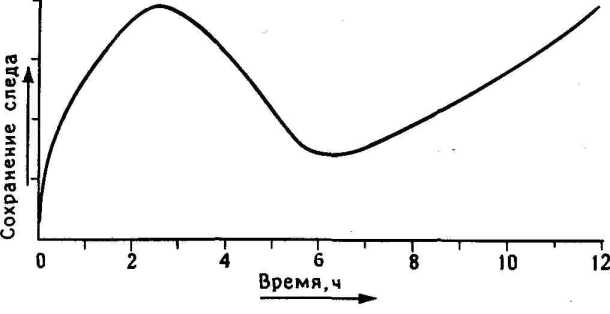
11.7. Двойная волна памяти. В этом опыте цыплятам в разное время до и после обучения (О) вводили ингибитор синтеза гликопротеинов 2-дГал и спустя 24 часа после тренировки тестировали память. Запоминание отсутствовало, если 2-дГал вводили в первые 12 часа после обучения или спустя 57 часов после него.
Амнестический эффект не наблюдался при введений 2-дГал в интервале между этими сроками или поздне 8 часов после обучения.
пробелы в знаниях, занимаясь биохимией. Если результаты опытов с повреждением мозга отражали реальные события, то вскоре после обучения должны были происходить биохимические изменения в левом IMHV, а позже такие же изменения должны появляться и в LPO. Значит, наряду с потоком информации из IMHV к LPO должен существовать "поток" биохимических изменений. Неожиданно я вспомнил о странных результатах, полученных магдебургской группой лет десять или больше назад в опытах на крысах, обучавшихся различению яркости. Я снова просмотрел статьи этих исследователей и убедился, что память не подвела меня.
Они наблюдали появление двух волн повышенного синтеза белков и, в частности, гликопротеинов: первой в гиппокампе, а второй, примерно через шесть часов после обучения, в коре мозга. В то время никто не знал, что можно извлечь из этих данных, и авторы не пошли дальше. В моей памяти начали всплывать разрозненные сведения, почерпнутые на недавних конференциях, которые в свое время, казалось, не представляли существенного интереса.
Как их проверить?
В нашей биохимической лаборатории мы еще изучали вопрос о гликопротеинах, используя ингибитор их синтеза 2-дГал. Работая ранее с Рейнхардом Йорком, я показал, что если вводить 2-дГал в мозг в любое время от двух часов до обучения до
двух часов после него, у цыплят развивается амнезия. Однако она не развивалась при инъекции спустя 3 часа после тренировки. Я не стал тогда изучать эффект введения 2-дГал в более поздние сроки, так как не видел в этом особого смысла, но теперь решил попробовать. Я повторил эксперимент, обучая цыплят по утрам, а затем вводя отдельным группам в разные сроки (с интервалом в 1 час) вплоть до 12 часов 2-дГал или для контроля физиологический раствор. На следующее утро, через 24 часа после обучения, я проверял их поведенческую реакцию.
Полученные результаты представлены на 11.7.
Первая часть кривой повторяет картину, выявленную раньше: инъекция 2-дГал вблизи периода обучения вызывала амнезию, а инъекция спустя 3 часа или позднее не давала эффекта. Это было так, пока я не подошел к шестичасовой отметке: тогда введение 2-дГал стало сопровождаться второй волной амнезии. Таким образом, в период после обучения должны быть две волны синтеза глйкопротеинов: одна сразу после обучения, а вторая спустя несколько часов. Не локализована ли первая волна в IMHV, а вторая в LPO?
Может быть, синтезируются разные гликопротеины? Это как раз те вопросы, что я изучал в эксперименте, который описан в главе 2, когда весной 1992 года начал писать эту книгу. Больше года нас осаждали бесчисленные технические проблемы. Я возненавидел эти проклятые гликопротеины и заодно себя за то, что вспомнил
0 двойной волне. Но я не собираюсь останавливаться на полпути,
хотя ответ на поставленные вопросы лишь смутно брезжит где-
то вдали. Сейчас, когда я пишу эти строки, выясняется, что
мы скорее всего правы: две волны различны, и вторая
локализована в LPO. Но время; ofnynjeHHW мне найсследб-"
вания, на исходе, и я опаздываю с представлением этой
рукописи. Если ко времени получения корректуры у меня
появятся новые данные, я постараюсь вставить их в книгу1.
Между тем нейрофизиологические исследования тоже шли полным ходом. Место Роджера Мейсона в паутине проводов и нагромождении осциллоскопов занял на этот раз с более сложным компьютерным оснащением очередной диссертант Джон Джигг, изучавший временную картину ритмической активности нейронов IMHV; одновременно он не упускал из
1 Я сейчас держу корректуру (май 1992 года) и на 90% уверен, что мы
правы.
318 1
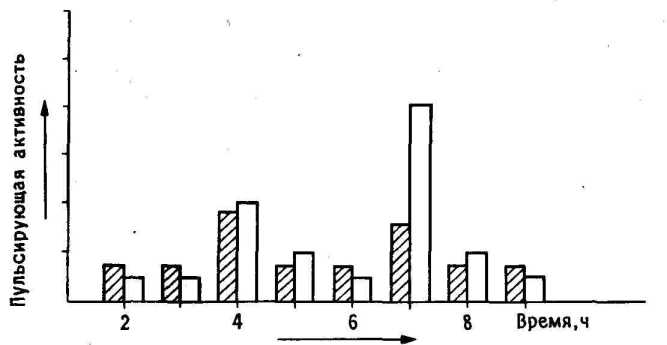
11.8. Динамика пульсирующей активности. На диаграмме показано усиление пульсирующей активности в левом (темные столбики) и правом (светлые' столбики) IMHV в различные сроки после обучения.
Обратите внимание, что спустя 34 часа после тренировки активность возрастала на обоих сторонах мозга, а спустя 67 часов она усиливалась в правом IMHV гораздо больше, чем в левом.
вида и LPO. Он получил необходимые данные примерно в то же время, когда мы пытались интерпретировать результаты повреждений мозга. Те и другие как будто согласовались между собой.
Сразу после обучения электрические разряды возникали в обоих полушариях, но спустя 46 часов их усиление регистрировалось главным образом в правом IMHV и одновременно они достигали максимальной интенсивности в правом, *г левом LPO ( 11.8).