
| 
|
|
|
|
Wallace и др.(1971) описывают более резкое и более быстрое, чем у контрольной группы, падение уровня лактата в крови во время ТМ с 1,24 моль/л (11,4 мг%) до 0,81 ммоль/л (7,3 мг%) при вышеуказанных значениях физического покоя.
Поскольку внутривенная инъекция лактата при отсутствии физической нагрузки, как известно, приводит к возникновению страха, то доказанное психологическими тестами снижение порогового уровня страха при практике ТМ может быть связано с этими данными (ср. Pitts 1969).
Гормоны коры надпочечников причисляются к «стрессовым гормонам» и это означает, что при т.н. «реакции борьбы и бегства» (см. 5.4.) их уровень в крови растет.
Jevning, Wilson и Davidson (1978) обнаружили у 15 занимающихся ТМ значительное понижение уровня кортизола примерно на 25% по сравнению с обычным уровнем кортизола в состоянии покоя, установив тем самым антистрессовое действие ТМ.
5.3.6. Электроэнцефалограмма и ее выражение при медитации
ЭЭГ, которую впервые в 1929 году зарегистрировал у человека Berger, будучи легко доступным у бодрствующего человека сигналом, оказалась с тех пор в центре исследований деятельности мозга. С одной стороны, метод электроэнцефалографии позволяет обнаружить строго воспроизводимые корреляции частотных характеристик ЭЭГ с состоянием бодрствования, и равным образом, корреляции пространственного распределения изменений частот с соответствующими патологическими состояниями, вследствие чего ЭЭГ прочно вошла в клиническую практику.
Однако, с другой стороны, какое-либо функциональное объяснение физиологической основы того или иного вида ЭЭГ до сих пор отсутствует (Gastaut 1974).
У человека выделяют четыре физиологически актуальных диапазона частот: д-ритм (14 - 30Гц), регистрируемый в состоянии бодрствования при направлении внимания вовне или же при умственной деятельности; б-ритм (8 - 13Гц) - в состоянии бодрствования при релаксации с закрытыми глазами; и-ритм (4 - 7Гц) и д-ритм (0,3 - 3Гц) (последние два ритма наступают во сне, а д-ритм - исключительно в стадии глубокого сна). Особенно интересен б-ритм, нейронное происхождение которого было отчасти установлено.
Речь идет о периодических пульсациях электрических потенциалов, отводимых одновременно на всем неокортексе, точнее об их отражении в мембранных потенциалах поверхностного дендритового слоя. Причиной этого являются возбуждающие и тормозящие постсинаптические потенциалы из разных мест пирамидных клеток. Периодичность задается из таламуса.
В таламусе находится генератор б-ритма, который в свою очередь подвержен влиянию ретикулярной формации ствола головного мозга (подробности см.: Creutzfeld 1974). Существование нейронных структур, порождающих б-ритм, лишь отчасти может объяснить генезис этого явления, но смысл его остается неясен. В ретикулярной формации находятся центры управления бодрствованием.
Нарастание активности (arousal) ведет к такому влиянию на таламус, что б-ритм в ЭЭГ исчезает и заменяется более частым ритмом меньшей амплитуды (д-ритм). Эту корковую реакцию активации называют также реакцией десинхронизации. Напротив, с нарастанием глубины сна начинает преобладать более медленный ритм. Структурную цепочку - ретикулярная формация-таламус-кора - называют восходящей ретикулярной системой.
По-видимому, некоторое несомненное влияние на ЭЭГ оказывает обмен веществ, так что «...все состояния, которые обусловлены замедлением мозгового обмена веществ или зависят от него, дают похожие изменения ЭЭГ (прирост выраженности амплитуды б-ритма и замедление его частоты при переходе в переднюю часть черепа)» (Gastaut 1974). Ingvar (1976) нашел типичный б-ритм более выраженным в затылочной, чем в лобной части.
При этом кровоснабжение лобной части мозга по сравнению с затылочной, судя по утилизации радиоактивно помеченной глюкозы, осуществляется более интенсивно.
У здорового человека в состоянии релаксации, по данным машинного анализа ЭЭГ, доминирует б-ритм с элементами в-ритма, причем доля б-ритма при одинаковом уровне бодрствования бывает различной (15% европейцев вообще не имеют б-ритма) и, в соответствии с вышеизложенным, будет обратно пропорционально величине обмена веществ. Однако все это еще не объясняет физиологического значения ЭЭГ, что побудило Gastaut (1974) придти к заключению, которое справедливо еще и сегодня, что «...эти волны представляют собой лишь общий побочный продукт жизнедеятельности коры головного мозга, присущий всем видам животных, но в особенности приматам». б-ритм, и, пожалуй, весь диапазон ЭЭГ сравним с трескотней пишущей машинки (или, скажем, компьютера), но это не печатный текст (и даже не вычислительные операции компьютера).
Все вышесказанное относится только к так называемой фоновой ЭЭГ-активности. Что же касается корковых ответов на раздражения, то здесь они рассматриваются отдельно в 5.3.8.
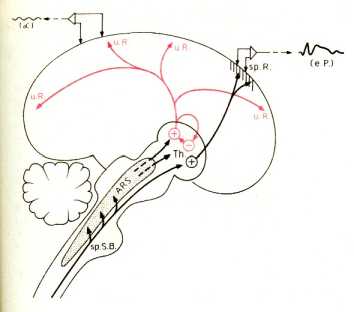
Они возбуждаются неспецифическими импульсами из восходящей ретикулярной системы (ARS). Чем сильнее эти импульсы, тем более высокие частоты обнаруживаются на ЭЭГ (активация). ARS возбуждается также и ответвлениями сенсорных путей, специфическая репрезентация (sp.R) которых в коре ведет к специфическим ощущениям и вызывает связанные с ним вызванные потенциалы (см., напр., 35). Во сне таламические задатчики ритма тормозятся влияниями т.н. центров сна в нижнем отделе ретикулярной формации, и на ЭЭГ регистрируются медленные ритмы. О функциональном назначении ритмов, генерируемых в таламусе ничего не известно (sp.
S.B. - специфический сенсорный путь; u.R. - неспецифическая репрезентация).
Какие выводы можно сделать из анализа ЭЭГ? Строго воспроизводимые корреляции позволяют по ритмам ЭЭГ у здорового человека однозначно установить степень вигильности, т.е. отличить состояние сна от бодрствования.
Разумеется, метод энцефалографии можно и следует применять для диагностики различных патологий. Однако анализ ЭЭГ позволяет делать лишь ограниченные выводы о некоторых психологических и психо-физиологических процессах.
В первой публикации по ЭЭГ при йогической медитации (Das и Gastaut 1955) описывается нарастающий во времени в-ритм, что отчасти противоречит более поздним публикациям. б-ритм, наличествующий лишь в затылочной доле, был ярко представлен до и после медитации, а во время медитации он сменялся на в-ритм, который из центральной области (Sulcus Rolandi - борозда Роланда) распространялся по всему кортексу. Из семи йогов один, по его собственному признанию, во время исследования достиг самадхи.
При этом амплитуда в-ритма (30-45Гц) у него достигла необычно высокой величины в 30-50 мкВ. Параллельно произошло легкое увеличение ЧСС и полное расслабление M.quadriceps femoris (четырехглавой мышцы бедра). По тем образцам ЭЭГ, которые представили Das и Gastaut, можно видеть последовательно изменения б-ритма в сторону более низких величин напряжения, его распространение в лобные доли черепа в начале медитации и далее вытеснение б-ритма все более сильно выраженным в-ритмом, который в итоге регистрировался на всем черепе.
Banquet (1972, 1973) также обнаружил это у некоторых из 10 практикующих ТМ посредством спектрального анализа и прямого просмотра ЭЭГ.
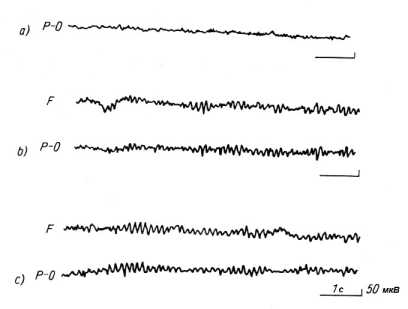
Аналогичные результаты незадолго до этого описали Bagсhi и Wenger (1957), причем подтверждения этому феномену даются всеми, кто занимался исследованиями йогических медитаций всякого рода (Rao и др. 1958, Hoenig 1968, Chhina 1974, Udupa и др. 1972, Dostбlek и др. 1979, 1980, Elson и др.
1977, Corby и др. 1978). б-ритм преобладает в каждом случае с замечательным постоянством.