
Функционирование сети APT в процессе классификации
Процесс классификации в APT состоит из трех основных фаз: распознавание, сравнение и поиск.Фаза распознавания.
В начальный момент времени входной вектор отсутствует на входе сети; следовательно, все компоненты входного вектора X можно рассматривать как нулевые. Тем самым сигнал G2 устанавливается в 0 и, следовательно, в нуль устанавливаются выходы всех нейронов слоя распознавания. Поскольку все нейроны слоя распознавания начинают работу в одинаковом состоянии, они имеют равные шансы выиграть в последующей конкуренции.
Затем на вход сети подается входной вектор X, который должен быть классифицирован. Этот вектор должен иметь одну или более компонент, отличных от нуля, в результате чего и G1, и G2 становятся равными единице. Это «подкачивает» нейроны слоя сравнения, обеспечивая один из двух единичных входов, необходимых для возбуждения нейронов в соответствии с правилом двух третей, тем самым позволяя нейрону возбуждаться, если соответствующая компонента входного вектора X равна единице. Таким образом, в течение данной фазы вектор S в точности дублирует вектор X.
Далее для каждого нейрона в слое распознавания вычисляется свертка вектора его весов Вj и вектора C (рис. 8.4). Нейрон с максимальным значением свертки имеет веса, наилучшим образом соответствующие входному вектору. Он выигрывает конкуренцию и возбуждается, одновременно затормаживая все остальные нейроны этого слоя. Таким образом, единственная компонента rj вектора R (рис. 8.1) становится равной единице, а все остальные компоненты становятся равными нулю.
В результате, сеть APT запоминает образы в весах нейронов слоя распознавания, один нейрон для каждой категории классификации. Нейрон слоя распознавания, веса которого наилучшим образом соответствуют входному вектору, возбуждается, его выход устанавливается в единичное значение, а выходы остальных нейронов этого слоя устанавливаются в нуль.
Фаза сравнения.
Единственный возбужденный в слое распознавания нейрон возвращает единицу обратно в слой сравнения в виде своего выходного сигнала rj. Эта единственная единица может быть визуально представлена в виде «веерного» выхода, подающегося через отдельную связь с весом tij на каждый нейрон в слое сравнения, обеспечивая каждый нейрон сигналом рj, равным величинеtij (нулю или единице) (рис. 8.5).
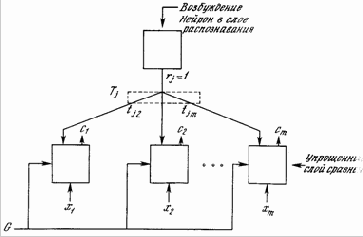
Рис. 8.5. Путь сигнала отдельного возбужденного нейрона в слое распознавания
Алгоритмы инициализации и обучения построены таким образом, что каждый весовой вектор Тj имеет двоичные значения весов; кроме того, каждый весовой вектор Вj представляет собой масштабированную версию соответствующего вектора Тj. Это означает, что все компоненты P (вектора возбуждения слоя сравнения) также являются двоичными величинами.
Так как вектор R не является больше нулевым, сигнал G1 устанавливается в нуль. Таким образом, в соответствии с правилом двух третей, возбудиться могут только нейроны, получающие на входе одновременно единицы от входного вектора X и вектора P.
Другими словами, обратная связь от распознающего слоя действует таким образом, чтобы установить компоненты C в нуль в случае, если входной вектор не соответствует входному образу, т. е. если X и P не имеют совпадающих компонент.
Если имеются существенные различия между X и P (малое количество совпадающих компонент векторов), несколько нейронов на фазе сравнения будут возбуждаться и C будет содержать много нулей, . в то время как X содержит единицы. Это означает, что возвращенный вектор P не является искомым и возбужденные нейроны в слое распознавания должны быть заторможены. Это торможение производится блоком сброса (рис. 8.1), который сравнивает входной вектор X и вектор C и вырабатывает сигнал сброса, если степень сходства этих векторов меньше некоторого уровня. Влияние сигнала сброса заключается в установке выхода возбужденного нейрона в нуль, отключая его на время текущей классификации.
Фаза поиска.
Если не выработан сигнал сброса, сходство является адекватным, и процесс классификации завершается. В противном случае другие запомненные образы должны быть исследованы с целью поиска лучшего соответствия. При этом торможение возбужденного нейрона в распознающем слое приводит к установке всех компонент вектора R в 0, G1 устанавливается в 1 и входной вектор X опять прикладывается в качестве C. В результате другой нейрон выигрывает соревнование в слое распознавания и другой запомненный образ P возвращается в слой сравнения. Если P не соответствует X, возбужденный нейрон в слое распознавания снова тормозится. Этот процесс повторяется до тех пор, пока не встретится одно из двух событий:
Найден запомненный образ, сходство которого с вектором X выше уровня параметра сходства, т. е. S>r. Если это происходит, проводится обучающий цикл, в процессе которого модифицируются веса векторов Tj и Bj, связанных с возбужденным нейроном в слое распознавания.
Все запомненные образы проверены, определено, что они не соответствуют входному вектору, и все нейроны слоя распознавания заторможены. В этом случае предварительно не распределенный нейрон в распознающем слое выделяется этому образу и его весовые векторы Bj и Tj устанавливаются соответствующими новому входному образу.
Проблема производительности.
Описанная сеть должна производить последовательный поиск среди всех запомненных образов. В аналоговых реализациях это будет происходить очень быстро; однако при моделировании на обычных цифровых компьютерах этот процесс может оказаться очень длительным. Если же сеть APT реализуется на параллельных процессорах, все свертки на распознающем уровне могут вычисляться одновременно. В этом случае поиск может быть очень быстрым.
Время, необходимое для стабилизации сети с латеральным торможением, может быть длительным при моделировании на последовательных цифровых компьютерах. Чтобы выбрать победителя в процессе латерального торможения, все нейроны в слое должны быть вовлечены в одновременные вычисления и передачу. Это может потребовать проведения большого объема вычислений перед достижением сходимости. Латеральные тормозящие сети, аналогичные используемым в неокогнитронах, могут существенно сократить это время (гл. 10).
Содержание раздела